

Protecting soil carbon is crucial for effective carbon management, as reversing losses is slow and difficult. This resource considers various effects on soil carbon in temperate managed ecosystems and can help land managers maintain and enhance carbon storage, supporting ecosystem resilience.
What is Soil Carbon? Why is it Important?
Soils store more carbon than the atmosphere and vegetation combined, an impressive fact that underscores the importance of considering soils when managing for ecosystem carbon. Not only do soils store vast amounts of carbon, but soil carbon also persists in the ecosystem for much longer than other pools of carbon. The mean residence time of soil carbon is on the order of decades to millennia, compared to vegetation carbon which cycles back to the atmosphere over timescales of years to centuries. The slow cycling of soil carbon also means accrual rates of new soil carbon are slow (Schlesinger 1990), while disturbance (e.g., land use change, erosion following biomass removal) can cause large and rapid site-level soil carbon losses (Guo and Gifford 2002, Berhe et al. 2018). Therefore, protecting existing soil carbon stores is foundational to managing for carbon because reversing soil carbon losses through management takes decades at a minimum, and is sometimes not even possible.
Soil organic carbon is primarily derived from plants, which convert atmospheric carbon dioxide (CO2 ) into organic matter via photosynthesis. Carbon initially fixed by plants and then consumed by other organisms such as animals and fungi also contributes to soil carbon. In non-arid systems, most soil carbon is organic carbon (hereafter, “soil carbon” refers to soil organic carbon) and is the focus of this primer. In contrast, arid systems have significant amounts of inorganic carbon in the form of calcium carbonate deposits, commonly referred to as “caliche”.
Plant organic matter is approximately 50% carbon, and a portion of this plant carbon is transferred to the soil. The remaining portion returns to the atmosphere as CO2 during plant respiration and microbial decomposition. There are three main pathways through which carbon moves from plants to soils: 1) exudation of carbon-rich molecules such as simple sugars from living plant roots into the soil, 2) decomposition of above- and below-ground dead plant tissue, and 3) transfer of carbon to mycorrhizal fungi (in exchange for resources such as nitrogen and phosphorus) which in turn is either released into soil by living fungi or incorporated into the soil as dead microbial biomass.
In general, carbon entering the soil from belowground roots or fungi contributes disproportionately to long-term soil carbon storage (Rasse et al. 2005, Jackson et al. 2017, Sokol and Bradford 2019). This is due in part to the proximity of belowground carbon inputs to soil minerals. Additionally, roots and fungi play large roles in promoting soil aggregation. Soil aggregation enhances soil carbon persistence because carbon occluded within aggregates is physically protected from microbial decomposition. Soil aggregation is hugely important to soil carbon storage and also contributes to soil water holding capacity and soil structure. Meanwhile, carbon from roots and fungi can also feed the microbial community, thereby accelerating microbial decomposition and loss of existing soil carbon. While this illustrates the complexity of soil carbon, overall, belowground inputs generally promote soil carbon storage (Verbruggen et al. 2013).
Carbon Storage
Soil Carbon Fractions
Carbon is stored in many different forms in the soil, affecting soil carbon sequestration and storage. To aid in characterizing and managing soil carbon, scientists commonly differentiate two broad fractions of soil carbon based on their distinct physical properties: particulate organic matter (POM), typically plantlike materials in various stages of decomposition ranging from recognizable plant debris to highly decomposed muck-like materials, and mineral associated organic matter (MAOM), which is mostly microbially-sourced organics that are preserved by bonding with mineral surfaces. These two procedurally defined soil fractions also vary in their process of formation, mean residence time, and response to climate change and land management (Cotrufo and Lavallee 2022), making the distinction between carbon in POM versus MAOM fractions useful for linking ecological processes with carbon management outcomes (Fig. 1)(Lavallee et al. 2020).
Carbon Sequestration
POM is formed by the physical transfer of plant, animal, and microbial products to the soil, while MAOM is formed from dissolved organic carbon (DOC) or microbial products adsorbing to soil mineral surfaces. Therefore, much of the carbon that initially enters the soil is in the form of POM, with some DOC leached or exuded from plant tissues directly entering the MAOM carbon pool. Over time, as POM is decomposed, a fraction of POM carbon is transferred to the MAOM pool while the remaining carbon is released into the atmosphere as CO2 . To illustrate, consider a deciduous leaf that falls to the ground in autumn. As the leaf slowly decomposes over weeks and months, carbon-rich leaf fragments will enter the POM pool. A proportion of carbon will quickly leach from the leaf and become DOC. Some DOC will move into the mineral soil and associate with soil minerals to become MAOM carbon, while the remaining DOC will leach out of the soils and drain into the watershed. Over time, the leaf fragments in the POM pool will be further decomposed by microbes, and this microbially-processed carbon will become part of the MAOM pool.
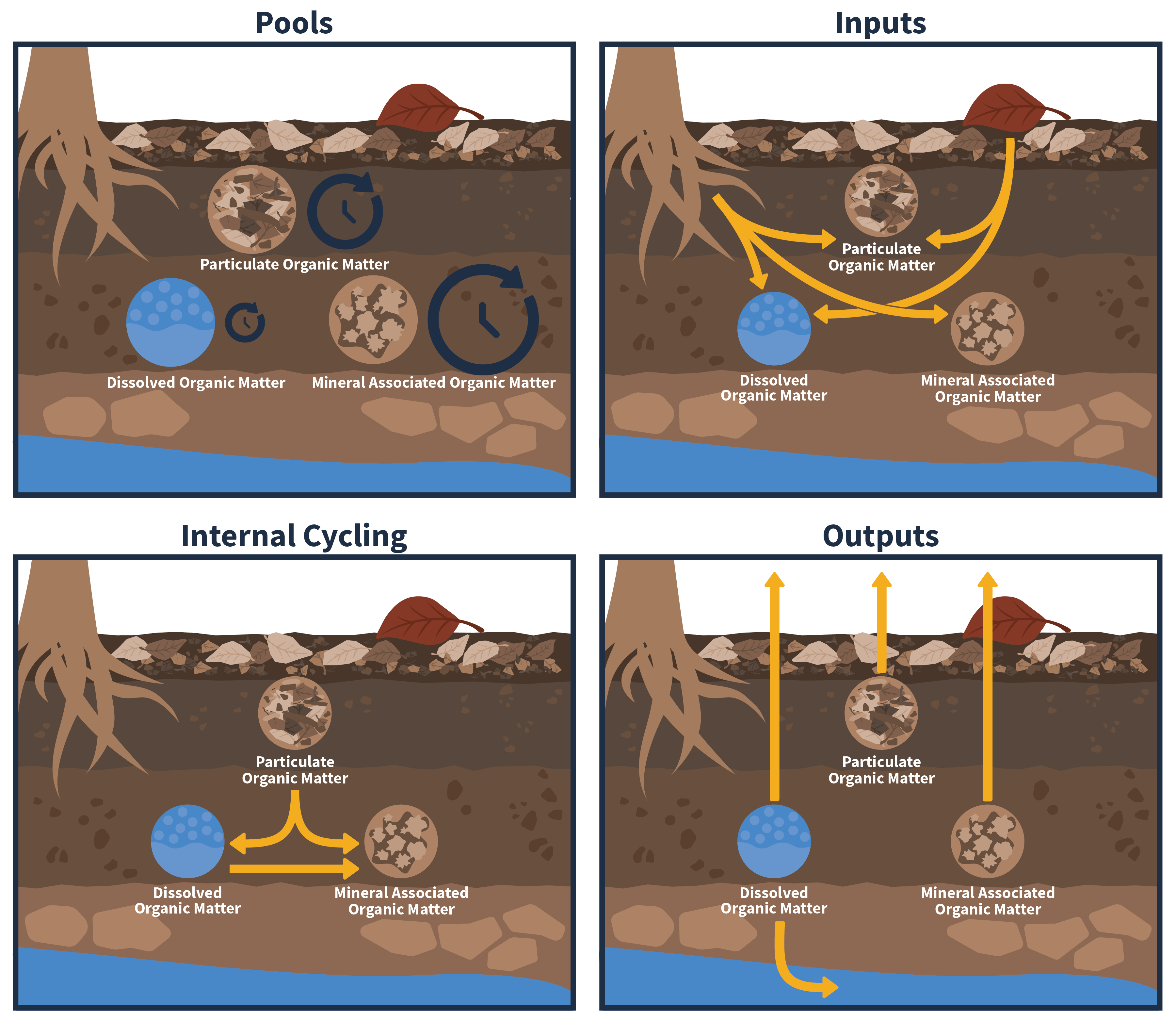
Historically, our understanding was that more complex carbon compounds (such as ‘tougher’ lignin-rich plant tissues) persist in the soil for longer. Now it is understood that carbon residence time is more influenced by protection from microbial processing (i.e., physically inaccessible to microbes or stuck strongly to soil mineral surfaces) than the chemical makeup of the organic matter. Inputs that decompose more quickly (e.g., fine roots) contribute more to MAOM carbon and long-term soil carbon storage compared to slower-decomposing carbon tissues such as downed woody debris (Cotrufo et al. 2013, 2015). Therefore, these relatively simple and fast-decomposing tissues still have a long residence time.
MAOM carbon is more resistant to carbon losses compared to POM due to binding with soil minerals, although POM contained in soil aggregates can also persist if soil structure is maintained. With increasing soil depth, relatively more carbon is held in MAOM and less in POM. As deeper soils are less prone to disturbance and decomposition, this pattern also lends to MAOM persistence. In general, MAOM has a longer mean residence time and therefore contributes more to long-term soil carbon storage compared to POM. Note that this distinction in residence time between MAOM and POM broadly holds true, but a significant portion of MAOM can turn over quickly.
Various Effects on Soil Carbon
Soil carbon storage is affected by many factors that interact over a wide range of spatial scales. As a result, spatial variation in soil carbon storage is large, which can make it challenging to know how much soil carbon is stored in a given place. Land managers can consider several important site factors to better understand a site’s relative current and potential soil carbon storage.
- Depth to bedrock: Deep soils will store more carbon on a per hectare basis compared to shallow soils, simply because there is more soil volume. Additionally, carbon stored deeper in soils generally persists for longer due to the dominance of MAOM over POM and less microbial decomposition and disturbance.
- Soil texture: Soils with high clay content (and, in particular, high concentrations of iron and aluminum oxides) have greater capacity to store carbon compared to sandy soils. Clays bind carbon to form MAOM, reducing leaching of carbon and promoting long-term persistence of soil carbon.
- Plant productivity and biomass: Plants are the dominant source of new carbon to soils, so sites with high plant productivity or biomass will have high potential for soil carbon storage. (Note that the persistence of these carbon inputs to soil will depend on factors such as soil texture, see above). Plant belowground carbon inputs, through roots and their associated mycorrhizal fungi, contribute disproportionately to persistent soil carbon and therefore plant species with high belowground carbon allocation (e.g., deep-rooted species, perennial grassland species, strong associations with mycorrhizal fungi) promote soil carbon.
- Soil microbial community: Soil microbes decompose organic matter. While this process releases some carbon back to the atmosphere, the process of decomposition makes the remaining carbon more persistent. Mycorrhizal fungi contribute significant amounts of carbon to the soil (Frey 2019, See et al. 2022) and play an important role in redistributing carbon into protected soil pools. Collectively, sites with a large, diverse, and active soil microbial community have greater potential for long-term soil carbon storage
Direct effects of climate change on soil carbon are harder to observe than climate change effects on vegetation. POM carbon pools, most prevalent in surface soils, are more sensitive to climate change because warmer and wetter conditions will accelerate decomposition. By contrast, the MAOM pools that dominate in deeper soils are less sensitive to climate, and are primarily controlled by soil properties such as texture and nitrogen availability to plants and microbes (Bai and Cotrufo 2022). The overall effects of future climate change on soil carbon sequestration and storage remain challenging to predict, with contrasting effects across sites likely due to the influence of local site conditions. Notably, recent increases in atmospheric CO2 have increased ecosystem carbon storage by increasing plant productivity (and subsequently increasing carbon inputs to soil). However, this ‘CO2 fertilization’ effect is expected to diminish as other resources (e.g., nutrients, water) limit plant productivity and ecosystem carbon cycling (Walker et al. 2021). Land managers can consider a few key site and soil characteristics to assess the potential effect of climate change on soil carbon:
- Are you managing a peatland? In peat soils, rising temperatures and lowered water tables (e.g., via ditching, tiling, or draining) will increase decomposition and drive a net loss in soil carbon. Rewetting peatlands leads to complex carbon cycling responses with the net effect on carbon sequestration and storage difficult to predict.
- Is climate change altering the fire regime at your site? Fire generally reduces forest floor organic soil carbon, with minimal effects on mineral soil carbon storage. These patterns vary depending on region and other site factors, with Pacific Northwest forests most likely to lose mineral soil carbon following fire. Forests that store more carbon in surface soils (i.e., thick organic horizons) are more vulnerable to soil carbon loss from fire. Wildfires, particularly high intensity fires that increase soil erosion, generally reduce soil carbon more than prescribed fires. Overall, fire decreases soil carbon storage in the short-term, with some of the remaining carbon transformed into more persistent forms. Over decades to centuries and barring additional major disturbances, soil carbon pools can recover to pre-fire levels (Nave et al. 2011, Pellegrini et al. 2021).
- Is your site prone to erosion? Climate change is resulting in more frequent extreme rain events, which may increase the likelihood of erosion at many sites, resulting in loss of surface soil carbon. Heavy erosion also exposes deeper soil carbon, decreasing its stability due to physical disturbance, increased weathering, and increased contact with decomposers
In most soils, it is not feasible to significantly increase soil carbon over management-relevant time scales. On the other hand, poor land management or land use change (e.g., clearing of forest for agriculture or development) can quickly deplete soil carbon stores. It is then useful to acknowledge several realities about spatial variation before considering soil carbon management. First, at landscape and larger scales, there is far more variation in soil carbon due to spatial variation in natural site factors than any degree of change that might result from management. Second, even within a site, spatial variation is usually larger than any changes that occur with management. From here, the key takeaway is that compared to natural variation, management effects tend to be small and generally restricted to surface soils. Below, general trends are summarized but site conditions are also critical factors to consider when predicting management effects on soil carbon.
- Harvesting and thinning: Harvesting in forests generally decreases forest floor (i.e., organic horizon) soil carbon, with greater losses in hardwood compared to mixed or coniferous stands. Harvest effects on mineral soil carbon storage is minimal, particularly in deeper soils (Nave et al. 2010).
- Prescribed fire: Fire combusts organic matter, releasing carbon into the atmosphere that may otherwise have been transferred to the soil. Prescribed fires generally reduce forest floor carbon. Effects of fire on mineral soils are weaker, with the greatest effects occurring with higher fire frequency and in sandy soils (Nave et al. 2011, Pellegrini et al. 2020). Prescribed fire may decrease carbon over the short term but can also decrease the risk of high intensity wildfires and associated carbon losses in the future.
Nave et al. (2021) synthesize management effects on soil carbon, with a focus on forests in the U.S. Lake States. As part of this synthesis, the researchers identified specific management tactics from the Menu of Adaptation and Mitigation Strategies and Approaches for Forest Carbon Management (Ontl et al. 2020) that could be used for carbon management goals. These tactics are meant to serve as examples, and different specific tactics will be appropriate for different sites. For each of the management categories bulleted above, select tactics are presented in Appendix 1 of this document, alongside a brief rationale connecting the tactic to one or more basic soil carbon principles presented here.
Appendix
| Management Category | Forest Carbon Menu Approach # | Tactic | Rationale for Tactic |
|---|---|---|---|
| Harvest | 2.1 | Conduct mechanized harvesting on vulnerable soil textures only during intermediate moisture, frozen (>15 cm), or snow-covered (>30 cm) conditions | Protect soil structure |
| Harvest | 2.1 | On soils with organic horizons, use slash mats and rubber-tired (not tracked) machines to preserve carbon-rich organic horizons and minimize mineral soil exposure; route trails parallel to slope gradients | Protect carbon-rich organic soil; prevent mineral soil exposure which increases mineral soil carbon decomposition and erosion |
| Harvest | 2.1 | Broadcast residues (if possible, chipped) on impacted sites such as forwarder trails, decking and land areas, and haul roads | Protect soil structure; reduce soil erosion; increase carbon inputs to soil from residues |
| Harvest | 2.1 | Wash adhering soil off of machines before moving from earthworm-invaded to uninvaded timber sales, but do not require washing for machines moving between uninvaded areas | Reduce decomposition from invasive earthworms, which eliminate organic horizons |
| Harvest | 2.1, 6.1 | During processing of individual trees, position residues (e.g., tops, limbs, reject logs) over areas of exposed mineral soil | Reduce soil carbon decomposition by mitigating increases in insolation and soil temperatures |
| Harvest | 2.1, 2.2, 5.1 | On wet sites, fell or redistribute residues in areas prone to saturation or ponding | Increase soil carbon inputs in low-oxygen environments where decomposition is slow |
| Harvest | 2.2 | Orient high basal area removal harvest patches with long axes running from E to W (rather than N to S) | Minimize decomposition by mitigating increases in insolation and soil temperatures |
| Harvest, Fire | 2.1, 2.2 | On sloping landforms, use contour felling and leave strips of residues or untreated forest. If slopes are compound, configure surface control features to connect, collect, and stabilize mobilized materials | Reduce soil erosion; increase carbon inputs to soil from residues |
| Fire | 3.2, 3.3 | In peatlands or upland-peatland complexes, prioritize wetland margins (especially on outwash) for wildfire suppression | Prevent permanent loss of wetland hydrology and carbonaccumulating conditions |
| Fire | 3.2, 3.3 | In peatlands, prioritize minerotrophic ecosystems rather than ombrotrophic ecosystems for wildfire suppression | Prevent permanent loss of wetland hydrology and carbonaccumulating conditions |
| Fire | 3.2, 3.3 | In uplands, prioritize deciduous rather than coniferous forests for wildfire suppression | Promote persistent forms of soil carbon (greater amounts of soil carbon are in stable pyrogenic forms in conifer vs. deciduous stands) |
| Restoration | 2.1 | Protect the soil surface during pine plantation harvesting if prescribed fire is to follow | Protect carbon-rich organic horizons from combined disturbance of harvest and fire |
| Restoration | 2.1 | In soils high in base cations, use harvest rather than fire (or harvest + fire) to restore early successional habitat | Promote persistent forms of soil carbon (base cations + fire can inhibit MAOM accumulation) |
| Restoration | 2.1 | In soils high in base cations, use harvest rather than fire (or harvest + fire) to restore early successional habitat | Promote persistent forms of soil carbon (base cations + fire can inhibit MAOM accumulation) |
| Restoration | 3.1, 3.3 | Use combination of mechanical treatments and low intensity prescribed fire to reduce fuels and fire risk in vulnerable landscapes | Reduce ecosystem vulnerability |
| Restoration | 6.2 | Limit number of stand entries for converting pine plantations to pine parklands or barrens | Protect soil structure |
| Restoration | 7.2 | Focus pine plantation to barrens restoration efforts on soils with the least surface organic matter, using summer burning | Target the restoration of low-carbon ecosystem types to sites that already have the least carbon |
Berhe, A. A.; Barnes, R. T. Six, J.; Marín-Spiotta, E. 2018. Role of Soil Erosion in Biogeochemical Cycling of Essential Elements: Carbon, Nitrogen, and Phosphorus. Annual Review of Earth and Planetary Sciences, 46: 521–548. doi. org/10.1146/annurev-earth-082517-010018
Cotrufo, M. F.; Lavallee, J. M. 2022. Soil organic matter formation, persistence, and functioning: A synthesis of current understanding to inform its conservation and regeneration. Advances in Agronomy, 172: 1–66. Elsevier. doi.org/10.1016/bs.agron.2021.11.002
Cotrufo, M. F.; Soong, J. L.; Horton, A. J.; Campbell, E. E.; Haddix, M. L.; Wall, D. H.; Parton, W. J. 2015. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nature Geoscience, 8: 776–779. doi.org/10.1038/ngeo2520
Cotrufo, M. F.; Wallenstein, M. D.; Boot, C. M.; Denef, K.; Paul, E. 2013. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Global Change Biology, 19: 988–995. doi.org/10.1111/gcb.12113.
Frey, S. D. 2019. Mycorrhizal Fungi as Mediators of Soil Organic Matter Dynamics. Annual Review of Ecology, Evolution, and Systematics, 50: 237–259. doi.org/10.1146/annurev-ecolsys-110617-062331.
Guo, L. B.; Gifford, R. M. 2002. Soil carbon stocks and land use change: a meta analysis. Global Change Biology, 8: 345–360. doi.org/10.1046/j.1354-1013.2002.00486.x
Jackson, R. B.; Lajtha, K.; Crow, S. E.; Hugelius, G.; Kramer, M. G.; Piñeiro, G. 2017. The Ecology of Soil Carbon: Pools, Vulnerabilities, and Biotic and Abiotic Controls. Annual Review of Ecology, Evolution, and Systematics, 48: 419– 445. doi.org/10.1146/annurev-ecolsys-112414-054234.
Lavallee, J. M.; Soong, J. L.; Cotrufo, M. F. 2020. Conceptualizing soil organic matter into particulate and mineral associated forms to address global change in the 21st century. Global Change Biology, 26: 261–273. doi. org/10.1111/gcb.14859
Nave, L. E.; DeLyser, K.; Domke, G. M.; Janowiak, M. K.; Ontl, T. A.; Sprague, E.; Walters, B. F.; C. W. Swanston. 2021. Land use and management effects on soil carbon in U.S. Lake States, with emphasis on forestry, fire, and reforestation. Ecological Applications 31, e02356. doi.org/10.1002/eap.2356.
Nave, L. E.; Vance, E. D.; Swanston, C. W.; Curtis, P. S. 2010. Harvest impacts on soil carbon storage in temperate forests. Forest Ecology and Management, 259: 857–866. doi.org/10.1016/j.foreco.2009.12.009.
Nave, L. E.; Vance, E. D.; Swanston, C. W.; Curtis, P. S. 2011. Fire effects on temperate forest soil C and N storage. Ecological Applications, 21: 1189–1201. doi.org/10.1890/10-0660.1.
Ontl, T. A.; Janowiak, M. K.; Swanston, C. W.; Daley, J.; Handler, S.; Cornett, M.; Hagenbuch, S.; Handrick, C.; Mccarthy, L.; Patch, N. 2020. Forest Management for Carbon Sequestration and Climate Adaptation. Journal of Forestry, 118: 86–101. doi.org/10.1093/jofore/fvz062.
Pellegrini, A. F. A.; Caprio, A. C.; Georgiou, K.; Finnegan, C.; Hobbie, S. E.; Hatten, J. A.; Jackson, R. B. 2021. Low intensity frequent fires in coniferous forests transform soil organic matter in ways that may offset ecosystem carbon losses. Global Change Biology, 27: 3810–3823. doi.org/10.1111/gcb.15648.
Pellegrini, A. F. A.; McLauchlan, K. K.; Hobbie, S. E.; Mack, M. C.; Marcotte, A. L.; Nelson, D. M.; Perakis, S. S.; Reich, P. B.; Whittinghill, K. 2020. Frequent burning causes large losses of carbon from deep soil layers in a temperate savanna. Journal of Ecology, 108: 1426–1441. doi.org/10.1111/1365-2745.13351.
Pellegrini, A. F. A.; McLauchlan, K. K.; Hobbie, S. E.; Mack, M. C.; Marcotte, A. L.; Nelson, D. M.; Perakis, S. S.; Reich, P. B.; Whittinghill, K. 2020. Frequent burning causes large losses of carbon from deep soil layers in a temperate savanna. Journal of Ecology, 108: 1426–1441. doi.org/10.1111/1365-2745.13351.
Rasse, D. P.; Rumpel, C.; Dignac, M.-F. 2005. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant and Soil, 269: 341–356. doi.org/10.1007/s11104-004-0907-y
See, C. R.; Keller, A. B.; Hobbie, S. E; Kennedy, P. G.; Weber, P. K.; Pett-Ridge, J. 2022. Hyphae move matter and microbes to mineral microsites: Integrating the hyphosphere into conceptual models of soil organic matter stabilization. Global Change Biology, 28: 2527–2540. doi.org/10.1111/gcb.16073.
Sokol, N. W.; Bradford, M. A. 2019. Microbial formation of stable soil carbon is more efficient from belowground than aboveground input. Nature Geoscience, 12: 46–53. doi.org/10.1038/s41561-018-0258-6
Verbruggen, E.; Veresoglou, S. D.; Anderson, I. C.; Caruso, T.; Hammer, E. C.; Kohler, J.; Rillig, M. C. 2013. Arbuscular mycorrhizal fungi – short-term liability but long-term benefits for soil carbon storage? New Phytologist, 197:,366–368. https://doi.org/10.1111/nph.12079.
Walker, A. P.; De Kauwe, M. G.; Bastos, A.; Belmecheri, S.; Georgiou, K.; Keeling, R.; McMahon, S. M.; Medlyn, B. E.; Moore, D. J. P.; Norby, R. J.; Zaehle, S.; Anderson-Teixeira, K. J.; Battipaglia, G.; Brienen, R. J. W.; Cabugao, K. G.; Cailleret, M.; Campbell, E.; Canadell, J.; Caias, P.; Craig, M. E.; Ellsworth, D. S.; Farquhar, G. D.; Fatichi, S.; Fisher, J. B.; Frank, D. C.; Graven, H; Gu, L.; Haverd, V.; Heilman, K.; Heimann, M.; Hungate, B. A.; Iversen, C. M.; Joos, F.; Jiang, M.; T. F. Keenan, T. F.; Knauer, J.; Körner, C.; Leshyk, V. O.; Leuzinger, S.; Liu, Y.; MacBean, N.; Malhi, Y.; McVicar, T. R.; Penuelas J.; Pongratz, J.; Powell, A. S.; Riutta, T.; Sabot, M. E. B.; Schleucher, J.; Sitch, S.; Smith, W. K.; Sulman, B.; Taylor, B.; Terrer, C.; Torn, M. S.; Treseder, K. K.; Trugman, A. T.; Trumbore, S. E.; van Mantgem, P. J.; Voelker, S. L.; Whelan, M. E.; Zuidema, P. A. 2021. Integrating the evidence for a terrestrial carbon sink caused by increasing atmospheric CO2 . New Phytologist, 229: 2413–2445. doi.org/10.1111/nph.16866.
Suggested Citation of the Hub Publication
Keller, A.B. and Handler, S. 2024. Soil organic carbon in temperate managed ecosystems: a primer. Technology Transfer. Houghton, MI: U.S. Department of Agriculture, Northern Forests Climate Hub. 8 p. https://doi.org/10.32747/2024.8633528.ch
Acknowledgments
This is a product of the USDA Northern Forests Climate Hub and the Northern Institute of Applied Climate Science, a collaborative, multi-institutional partnership led by the USDA Forest Service. Funding was provided by the USDA Forest Service and The Nature Conservancy.